| Henry Gray (1821–1865). Anatomy of the Human Body. 1918. |
| |
| 2. Development of the Nervous System |
| |
| The entire nervous system is of ectodermal origin, and its first rudiment is seen in the neural groove which extends along the dorsal aspect of the embryo (Fig. 17). By the elevation and ultimate fusion of the neural folds, the groove is converted into the neural tube (Fig. 19). The anterior end of the neural tube becomes expanded to form the three primary brain-vesicles; the cavity of the tube is subsequently modified to form the ventricular cavities of the brain, and the central canal of the medulla spinalis; from the wall the nervous elements and the neuroglia of the brain and medulla spinalis are developed. | 1 |
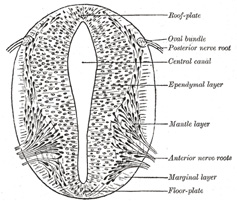 |
FIG. 640– Section of medulla spinalis of a four weeks’ embryo. (His.) (See enlarged image) | | |
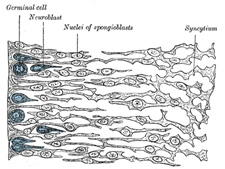 |
FIG. 641– Transverse section of the medulla spinalis of a human embryo at the beginning of the fourth week. The left edge of the figure corresponds to the lining of the central canal. (His.) (See enlarged image) | | |
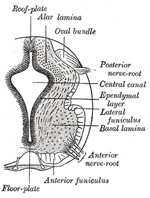 |
FIG. 642– aged about four and a half weeks. (See enlarged image) | | |
 |
FIG. 643– aged about three months. (See enlarged image) | | |
| | | The Medulla Spinalis.—At first the wall of the neural tube is composed of a single layer of columnar ectodermal cells. Soon the side-walls become thickened, while the dorsal and ventral parts remain thin, and are named the roof- and floor-plates (Figs. 640, 642, 643). A transverse section of the tube at this stage presents an oval outline, while its lumen has the appearance of a slit. The cells which constitute the wall of the tube proliferate rapidly, lose their cell-boundaries and form a syncytium. This syncytium consists at first of dense protoplasm with closely packed nuclei, but later it opens out and forms a looser meshwork with the cellular strands arranged in a radiating manner from the central canal. Three layers may now be defined—an internal or ependymal, an intermediate or mantle, and an external or marginal. The ependymal layer is ultimately converted into the ependyma of the central canal; the processes of its cells pass outward toward the periphery of the medulla spinalis. The marginal layer is devoid of nuclei, and later forms the supporting framework for the white funiculi of the medulla spinalis. The mantle layer represents the whole of the future gray columns of the medulla spinalis; in it the cells are differentiated into two sets, viz., (a) spongioblasts or young neuroglia cells, and (b) germinal cells, which are the parents of the neuroblasts or young nerve cells (Fig. 641). The spongioblasts are at first connected to one another by filaments of the syncytium; in these, fibrils are developed, so that as the neuroglial cells become defined they exhibit their characteristic mature appearance with multiple processes proceeding from each cell. The germinal cells are large, round or oval, and first make their appearance between the ependymal cells on the sides of the central canal. They increase rapidly in number, so that by the fourth week they form an almost continuous layer on each side of the tube. No germinal cells are found in the roof- or floor-plates; the roof-plate retains, in certain regions of the brain, its epithelial character; elsewhere, its cells become spongioblasts. By subdivision the germinal cells give rise to the neuroblasts or young nerve cells, which migrate outward from the sides of the central canal into the mantle layer and neural crest, and at the same time become pear-shaped; the tapering part of the cell undergoes still further elongation, and forms the axiscylinder of the cell. | 2 |
| The lateral walls of the medulla spinalis continue to increase in thickness, and the canal widens out near its dorsal extremity, and assumes a somewhat lozengeshaped appearance. The widest part of the canal serves to subdivide the lateral wall of the neural tube into a dorsal or alar, and a ventral or basal lamina (Figs. 642, 643), a subdivision which extends forward into the brain. At a later stage the ventral part of the canal widens out, while the dorsal part is first reduced to a mere slit and then becomes obliterated by the approximation and fusion of its walls; the ventral part of the canal persists and forms the central canal of the adult medulla spinalis. The caudal end of the canal exhibits a conical expansion which is known as the terminal ventricle. | 3 |
| The ventral part of the mantle layer becomes thickened, and on cross-section appears as a triangular patch between the marginal and ependymal layers. This thickening is the rudiment of the anterior column of gray substance, and contains many neuroblasts, the axis-cylinders of which pass out through the marginal layer and form the anterior roots of the spinal nerves (Figs. 640, 642, 643). The thickening of the mantle layer gradually extends in a dorsal direction, and forms the posterior column of gray substance. The axons of many of the neuroblasts in the alar lamina run forward, and cross in the floor-plate to the opposite side of the medulla spinalis; these form the rudiment of the anterior white commissure. | 4 |
| About the end of the fourth week nerve fibers begin to appear in the marginal layer. The first to develop are the short intersegmental fibers from the neuroblasts in the mantle zone, and the fibers of the dorsal nerve roots which grow into the medulla spinalis from the cells of the spinal ganglia. By the sixth week these dorsal root fibers form a well-defined oval bundle in the peripheral part of the alar lamina; this \??\ gradually increases in size, and spreading toward the middle line forms the rudiment of the posterior funiculus. The long intersegmental fibers begin to appear about the third month and the cerebrospinal fibers about the fifth month. All nerve fibers are at first destitute of medullary sheaths. Different groups of fibers receive their sheaths at different times—the dorsal and ventral nerve roots about the fifth month, the cerebrospinal fibers after the ninth month. | 5 |
| By the growth of the anterior columns of gray substance, and by the increase in size of the anterior funiculi, a furrow is formed between the lateral halves of the cord anteriorly; this gradually deepens to form the anterior median fissure. The mode of formation of the posterior septum is somewhat uncertain. Many believe that it is produced by the growing together of the walls of the posterior part of the central canal and by the development from its ependymal cells of a septum of fibrillated tissue which separates the future funiculi graciles. | 6 |
| Up to the third month of fetal life the medulla spinalis occupies the entire length of the vertebral canal, and the spinal nerves pass outward at right angles to the medulla spinalis. From this time onward, the vertebral column grows more rapidly than the medulla spinalis, and the latter, being fixed above through its continuity with the brain, gradually assumes a higher position within the canal. By the sixth month its lower end reaches only as far as the upper end of the sacrum; at birth it is on a level with the third lumbar vertebra, and in the adult with the lower border of the first or upper border of the second lumbar vertebra. A delicate filament, the filum terminale, extends from its lower end as far as the coccyx. | 7 |
| | | The Spinal Nerves.—Each spinal nerve is attached to the medulla spinalis by an anterior or ventral and a posterior or dorsal root. | 8 |
| The fibers of the anterior roots are formed by the axons of the neuroblasts which lie in the ventral part of the mantle layer; these axons grow out through the overlying marginal layer and become grouped to form the anterior nerve root (Fig. 641). | 9 |
| The fibers of the posterior roots are developed from the cells of the spinal ganglia. Before the neural groove is closed to form the neural tube a ridge of ectodermal cells, the ganglion ridge or neural crest (Fig. 644), appears along the prominent margin of each neural fold. When the folds meet in the middle line the two ganglion ridges fuse and form a wedge-shaped area along the line of closure of the tube. The cells of this area proliferate rapidly opposite the primitive segments and then migrate in a lateral and ventral direction to the sides of the neural tube, where they ultimately form a series of oval-shaped masses, the future spinal ganglia. These ganglia are arranged symmetrically on the two sides of the neural tube and, except in the region of the tail, are equal in number to the primitive segments. The cells of the ganglia, like the cells of the mantle layer, are of two kinds, viz., spongioblasts and neuroblasts. The spongioblasts develop into the neuroglial cells of the ganglia. The neuroblasts are at first round or oval in shape, but soon assume the form of spindles the extremities of which gradually elongate into central and peripheral processes. The central processes grow medialward and, becoming connected with the neural tube, constitute the fibers of the posterior nerve roots, while the peripheral processes grow lateralward to mingle with the fibers of the anterior root in the spinal nerve. As development proceeds the original bipolar form of the cells changes; the two processes become approximated until they ultimately arise from a single stem in a T-shaped manner. Only in the ganglia of the acoustic nerve is the bipolar form retained. More recent observers hold, however, that the T-form is derived from the branching of a single process which grows out from the cell. | 10 |
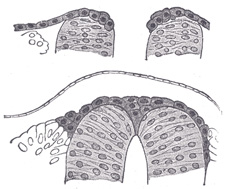 |
FIG. 644– Two stages in the development of the neural crest in the human embryo. (Lenhossèk.) (See enlarged image) | | |
| The anterior or ventral and the posterior or dorsal nerve roots join immediately beyond the spinal ganglion to form the spinal nerve, which then divides into anterior, posterior, and visceral divisions. The anterior and posterior divisions proceed directly to their areas of distribution without further association with ganglion cells (Fig. 645). The visceral divisions are distributed to the thoracic, abdominal, and pelvic viscera, to reach which they pass through the sympathetic trunk, and many of the fibers form arborizations around the ganglion cells of this trunk. Visceral branches are not given off from all the spinal nerves; they form two groups, viz., (a) thoracico-lumbar, from the first or second thoracic, to the second or third lumbar nerves; and (b) pelvic, from the second and third, or third and fourth sacral nerves. | 11 |
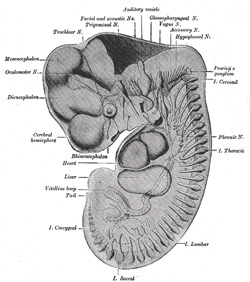 |
FIG. 645– Reconstruction of periphera nerves of a human embryo of 10.2 mm. (After His.) The abducent nerve is not labelled, but is seen passing forward to the eye under the mandibular and maxillary nerves. (See enlarged image) | | |
| | | The Brain.—The brain is developed from the anterior end of the neural tube, which at an early period becomes expanded into three vesicles, the primary cerebral vesicles (Fig. 18). These are marked off from each other by intervening constrictions, and are named the fore-brain or prosencephalon, the mid-brain or mesencephalon, and the hind-brain or rhombencephalon—the last being continuous with the medulla spinalis. As the result of unequal growth of these different parts three flexures are formed and the embryonic brain becomes bent on itself in a somewhat zigzag fashion; the two earliest flexures are concave ventrally and are associated with corresponding flexures of the whole head. The first flexure appears in the region of the mid-brain, and is named the ventral cephalic flexure (Fig. 650). By means of it the fore-brain is bent in a ventral direction around the anterior end of the notochord and fore-gut, with the result that the floor of the fore-brain comes to lie almost parallel with that of the hind-brain. This flexure causes the mid-brain to become, for a time, the most prominent part of the brain, since its dorsal surface corresponds with the convexity of the curve. The second bend appears at the junction of the hind-brain and medulla spinalis. This is termed the cervical flexure (Fig. 652), and increases from the third to the end of the fifth week, when the hind-brain forms nearly a right angle with the medulla spinalis; after the fifth week erection of the head takes place and the cervical flexure diminishes and disappears. The third bend is named the pontine flexure (Fig. 652), because it is found in the region of the future pons Varoli. It differs from the other two in that (a) its convexity is forward, and (b) it does not affect the head. The lateral walls of the brain-tube, like those of the medulla spinalis, are divided by internal furrows into alar or dorsal and basal or ventral laminæ (Fig. 646). | 12 |
 |
FIG. 646– Diagram to illustrate the alar and basal laminæ of brain vesicles. (His.) (See enlarged image) | | |
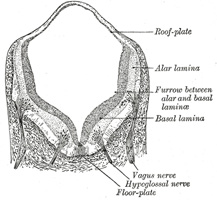 |
FIG. 647– Transverse section of medulla oblongata of human embryo. X 32. (Kollmann.) (See enlarged image) | | |
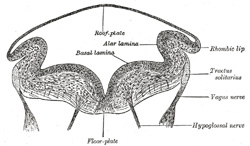 |
FIG. 648– Transverse section of medulla oblongata of human embryo. (After His.) (See enlarged image) | | |
| | | The Hind-brain or Rhombencephalon.—The cavity of the hind-brain becomes the fourth ventricle. At the time when the ventral cephalic flexure makes its appearance, the length of the hind-brain exceeds the combined lengths of the other two vesicles. Immediately behind the mid-brain it exhibits a marked constriction, the isthmus rhombencephali (Fig. 650, Isthmus), which is best seen when the brain is viewed from the dorsal aspect. From the isthmus the anterior medullary velum and the superior peduncle of the cerebellum are formed. It is customary to divide the rest of the hind-brain into two parts, viz., an upper, called the metencephalon, and a lower, the myelencephalon. The cerebellum is developed by a thickening of the roof, and the pons by a thickening in the floor and lateral walls of the metencephalon. The floor and lateral walls of the myelencephalon are thickened to form the medulla oblongata; its roof remains thin, and, retaining to a great extent its epithelial nature, is expanded in a lateral direction. Later, by the growth and backward extension of the cerebellum, the roof is folded inward toward the cavity of the fourth ventricle; it assists in completing the dorsal wall of this cavity, and is also invaginated to form the ependymal covering of its choroid plexuses. Above it is continuous with the posterior medullary velum; below, with the obex and ligulæ. | 13 |
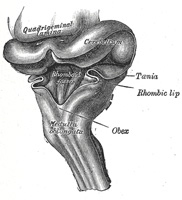 |
FIG. 649– Hind-brain of a human embryo of three months—viewed from behind and partly from left side. (From model by His.) (See enlarged image) | | |
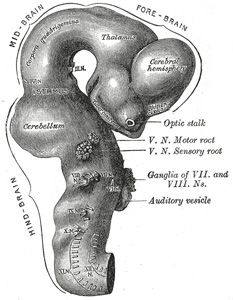 |
FIG. 650– Exterior of brain of human embryo of four and a half weeks. (From model by His.) (See enlarged image) | | |
| The development of the medulla oblongata resembles that of the medulla spinalis, but at the same time exhibits one or two interesting modifications. On transverse section the myelencephalon at an early stage is seen to consist of two lateral walls, connected across the middle line by floor- and roof-plates (Figs. 647 and 648). Each lateral wall consists of an alar and a basal lamina, separated by an internal furrow, the remains of which are represented in the adult brain by the sulcus limitans on the rhomboid fossa. The contained cavity is more or less triangular in outline, the base being formed by the roof-plate, which is thin and greatly expanded transversely. Pear-shaped neuroblasts are developed in the alar and basal laminæ. and their narrow stalks are elongated to form the axis-cylinders of the nerve fibers. Opposite the furrow or boundary between the alar and basal laminæ a bundle of nerve fibers attaches itself to the outer surface of the alar lamina. This is named the tractus solitarius (Fig. 648), and is formed by the sensory fibers of the glossopharyngeal and vagus nerves. It is the homologue of the oval bundle seen in the medulla spinalis, and, like it, is developed by an ingrowth of fibers from the ganglia of the neural crest. At first it is applied to the outer surface of the alar lamina, but it soon becomes buried, owing to the growth over it of the neighboring parts. By the fifth week the dorsal part of the alar lamina bends in a lateral direction along its entire length, to form what is termed the rhombic lip (Figs. 648, 649). Within a few days this lip becomes applied to, and unites with, the outer surface of the main part of the alar lamina, and so covers in the tractus solitarius and also the spinal root of the trigeminal nerve; the nodulus and flocculus of the cerebellum are developed from the rhombic lip. | 14 |
| Neuroblasts accumulate in the mantle layer; those in the basal lamina correspond with the cells in the anterior gray column of the medulla spinalis, and, like them, give origin to motor nerve fibers; in the medulla oblongata they are, however, arranged in groups or nuclei, instead of forming a continuous column. From the alar lamina and its rhombic lip, neuroblasts migrate into the basal lamina, and become aggregated to form the olivary nuclei, while many send their axis-cylinders through the floor-plate to the opposite side, and thus constitute the rudiment of the raphé of the medulla oblongata. By means of this thickening of the ventral portion, the motor nuclei are buried deeply in the interior, and, in the adult, are found close to the rhomboid fossa. This is still further accentuated: (a) by the development of the pyramids, which are formed about the fourth month by the downward growth of the motor fibers from the cerebral cortex; and (b) by the fibers which pass to and from the cerebellum. On the rhomboid fossa a series of six temporary furrows appears; these are termed the rhombic grooves. They bear a definite relationship to certain of the cranial nerves; thus, from before backward the first and second grooves overlie the nucleus of the trigeminal; the third, the nucleus of the facial; the fourth, that of the abducent; the fifth, that of the glossopharyngeal; and the sixth, that of the vagus. | 15 |
| The pons is developed from the ventro-lateral wall of the metencephalon by a process similar to that which has been described for the medulla oblongata. | 16 |
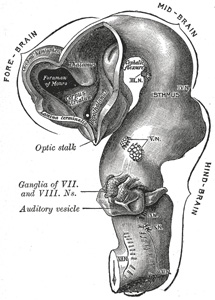 |
FIG. 651– Brain of human embryo of four and a half weeks, showing interior of fore-brain. (From model by His.) (See enlarged image) | | |
| The cerebellum is developed in the roof of the anterior part of the hind-brain (Figs. 649 to 654). The alar laminæ of this region become thickened to form two lateral plates which soon fuse in the middle line and produce a thick lamina which roofs in the upper part of the cavity of the hind-brain vesicle; this constitutes the rudiment of the cerebellum, the outer surface of which is originally smooth and convex. The fissures of the cerebellum appear first in the vermis and floccular region, and traces of them are found during the third month; the fissures on the cerebellar hemispheres do not appear until the fifth month. The primitive fissures are not developed in the order of their relative size in the adult—thus the horizontal sulcus in the fifth month is merely a shallow groove. The best marked of the early fissures are: (a) the fissura prima between the developing culmen and declive, and (b) the fissura secunda between the future pyramid and uvula. The flocculus and nodule are developed from the rhombic lip, and are therefore recognizable as separate portions before any of the other cerebellar lobules. The groove produced by the bending over of the rhombic lip is here known as the floccular fissure; when the two lateral walls fuse, the right and left floccular fissures join in the middle line and their central part becomes the post-nodular fissure. | 17 |
| On the ventricular surface of the cerebellar lamina a transverse furrow, the incisura fastigii, appears, and deepens to form the tent-like recess of the roof of the fourth ventricle. The rudiment of the cerebellum at first projects in a dorsal direction; but, by the backward growth of the cerebrum, it is folded downward and somewhat flattened, and the thin roof-plate of the fourth ventricle, originally continuous with the posterior border of the cerebellum, is projected inward toward the cavity of the ventricle. | 18 |
| | | The Mid-brain or Mesencephalon.—The mid-brain (Figs. 650 to 654) exists for a time as a thin-walled cavity of some size, and is separated from the isthmus rhombencephali behind, and from the fore-brain in front, by slight constrictions. Its cavity becomes relatively reduced in diameter, and forms the cerebral aqueduct of the adult brain. Its basal laminæ increase in thickness to form the cerebral peduncles, which are at first of small size, but rapidly enlarge after the fourth month. The neuroblasts of these laminæ are grouped in relation to the sides and floor of the cerebral aqueduct, and constitute the nuclei of the oculomotor and trochlear nerves, and of the mesencephalic root of the trigeminal nerve. By a similar thickening process its alar laminæ are developed into the quadrigeminal lamina. The dorsal part of the wall for a time undergoes expansion, and presents an internal median furrow and a corresponding external ridge; these, however, disappear, and the latter is replaced by a groove. Subsequently two oblique furrows extend medialward and backward, and the thickened lamina is thus subdivided into the superior and inferior colliculi. | 19 |
 |
FIG. 652– Exterior of brain of human embryo of five weeks. (From model by His.) (See enlarged image) | | |
| | | The Fore-brain or Prosencephalon.—A transverse section of the early fore-brain shows the same parts as are displayed in similar sections of the medulla spinalis and medulla oblongata, viz., a pair of thick lateral walls connected by thin floor-and roof-plates. Moreover, each lateral wall exhibits a division into a dorsal or alar and a ventral or basal lamina separated internally by a furrow termed the sulcus of Monro. This sulcus ends anteriorly at the medial end of the optic stalk, and in the adult brain is retained as a slight groove extending backward from the interventricular foramen to the cerebral aqueduct. | 20 |
| At a very early period—in some animals before the closure of the cranial part of the neural tube—two lateral diverticula, the optic vesicles, appear, one on either side of the fore-brain; for a time they communicate with the cavity of the fore-brain by relatively wide openings. The peripheral parts of the vesicles expand, while the proximal parts are reduced to tubular stalks, the optic stalks. The optic vesicle gives rise to the retina and the epithelium on the back of the ciliary body and iris; the optic stalk is invaded by nerve fibers to form the optic nerve. The fore-brain then grows forward, and from the alar laminæ of this front portion the cerebral hemispheres originate as diverticula which rapidly expand to form two large pouches, one on either side. The cavities of these diverticula are the rudiments of the lateral ventricles; they communicate with the median part of the fore-brain cavity by relatively wide openings, which ultimately form the interventricular foramen. The median portion of the wall of the fore-brain vesicle consists of a thin lamina, the lamina terminalis (Figs. 654, 657), which stretches from the interventricular foramen to the recess at the base of the optic stalk. The anterior part of the fore-brain, including the rudiments of the cerebral hemispheres, is named the telencephalon, and its posterior portion is termed the diencephalon; both of these contribute to the formation of the third ventricle. | 21 |
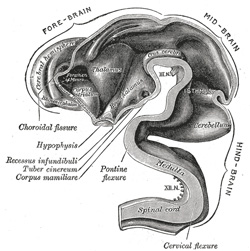 |
FIG. 653– Interior of brain of human embryo of five weeks. (From model by His.) (See enlarged image) | | |
| | | The Diencephalon.—From the alar lamina of the diencephalon, the thalamus, metathalamus, and epithalamus are developed. The thalamus (Figs. 650 to 654) arises as a thickening which involves the anterior two-thirds of the alar lamina. The two thalami are visible, for a time, on the surface of the brain, but are subsequently hidden by the cerebral hemispheres which grow backward over them. The thalami extend medialward and gradually narrow the cavity between them into a slit-like aperture which forms the greater part of the third ventricle; their medial surfaces ultimately adhere, in part, to each other, and the intermediate mass of the ventricle is developed across the area of contact. The metathalamus comprises the geniculate bodies which originate as slight outward bulgings of the alar lamina. In the adult the lateral geniculate body appears as an eminence on the lateral part of the posterior end of the thalamus, while the medial is situated on the lateral aspect of the mid-brain. The epithalamus includes the pineal body, the posterior commissure, and the trigonum habenulæ. The pineal body arises as an upward the evagination of roof-plate immediately in front of the midbrian; this evagination becomes solid with the exception of its proximal part, which persists as the recessus pinealis. In lizards the pineal evagination is elongated into a stalk, and its peripheral extremity is expanded into a vesicle, in which a rudimentary lens and retina are formed; the stalk becomes solid and nerve fibers make their appearance in it, so that in these animals the pineal body forms a rudimentary eye. The posterior commissure is formed by the ingrowth of fibers into the depression behind and below the pineal evagination, and the trigonum habenulæ is developed in front of the pineal recess. | 22 |
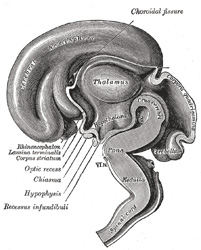 |
FIG. 654– Median sagittal section of brain of human embryo of three months. (From model by His.) (See enlarged image) | | |
| From the basal laminæ of the diencephalon the pars mamillaris hypothalami is developed; this comprises the corpora mamillaria and the posterior part of the tuber cinereum. The corpora mamillaria arise as a single thickening, which becomes divided into two by a median furrow during the third month. | 23 |
| The roof-plate of the diencephalon, in front of the pineal body, remains thin and epithelial in character, and is subsequently invaginated by the choroid plexuses of the third ventricle. | 24 |
| | | The Telencephalon.—This consists of a median portion and two lateral diverticula. The median portion forms the anterior part of the cavity of the third ventricle, and is closed below and in front by the lamina terminalis. The lateral diverticula consist of outward pouchings of the alar laminæ; the cavities represent the lateral ventricles, and their walls become thickened to form the nervous matter of the cerebral hemispheres. The roof-plate of the telencephalon remains thin, and is continuous in front with the lamina terminalis and behind with the roof-plate of the diencephalon. In the basal laminæ and floor-plate the pars optica hypothalami is developed; this comprises the anterior part of the tuber cinereum, the infundibulum and posterior lobe of the hypophysis, and the optic chiasma. The anterior part of the tuber cinereum is derived from the posterior part of the floor of the telencephalon; the infundibulum and posterior lobe of the hypophysis arise as a downward diverticulum from the floor. The most dependent part of the diverticulum becomes solid and forms the posterior lobe of the hypophysis; the anterior lobe of the hypophysis is developed from a diverticulum of the ectodermal lining of the stomodeum. The optic chiasma is formed by the meeting and partial decussation of the optic nerves, which subsequently grow backward as the optic tracts and end in the diencephalon. | 25 |
| The cerebral hemispheres arise as diverticula of the alar laminæ of the telencephalon (Figs. 650 to 654); they increase rapidly in size and ultimately overlap the structures developed from the mid- and hind-brains. This great expansion of the hemispheres is a characteristic feature of the brains of mammals, and attains its maximum development in the brain of man. Elliott-Smith divides each cerebral hemisphere into three fundamental parts, viz., the rhinencephalon, the corpus striatum, and the neopallium. | 26 |
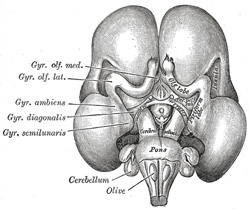 |
FIG. 655– Inferior surface of brain of embryo at beginning of fourth month. (From Kollmann.) (See enlarged image) | | |
| The rhinencephalon (Fig. 655) represents the oldest part of the telencephalon, and forms almost the whole of the hemisphere in fishes, amphibians, and reptiles. In man it is feebly developed in comparison with the rest of the hemisphere, and comprises the following parts, viz., the olfactory lobe (consisting of the olfactory tract and bulb and the trigonum olfactorium), the anterior perforated substance, the septum pellucidum, the subcallosal, supracallosal, and dentate gyri, the fornix, the hippocampus, and the uncus. The rhinencephalon appears as a longitudinal elevation, with a corresponding internal furrow, on the under surface of the hemisphere close to the lamina terminalis; it is separated from the lateral surface of the hemisphere by a furrow, the external rhinal fissure, and is continuous behind with that part of the hemisphere, which will ultimately form the anterior end of the temporal lobe. The elevation becomes divided by a groove into an anterior and a posterior part. The anterior grows forward as a hollow stalk the lumen of which is continuous with the anterior part of the ventricular cavity. During the third month the stalk becomes solid and forms the rudiment of the olfactory bulb and tract; a strand of gelatinous tissue in the interior of the bulb indicates the position of the original cavity. From the posterior part the anterior perforated substance and the pyriform lobe are developed; at the beginning of the fourth month the latter forms a curved elevation continuous behind with the medial surface of the temporal lobe, and consisting, from before backward, of the gyrus olfactorius lateralis, gyrus ambiens, and gyrus semilunaris, parts which in the adult brain are represented by the lateral root of the olfactory tract and the uncus. The position and connections of the remaining portions of the rhinencephalon are described with the anatomy of the brain. | 27 |
| The corpus striatum (Figs. 651 and 653) appears in the fourth week as a triangular thickening of the floor of the telencephalon between the optic recess and the interventricular foramen, and continuous behind with the thalamic part of the diencephalon. It increases in size, and by the second month is seen as a swelling in the floor of the future lateral ventricle; this swelling reaches as far as the posterior end of the primitive hemisphere, and when this part of the hemisphere grows backward and downward to form the temporal lobe, the posterior part of the corpus striatum is carried into the roof of the inferior horn of the ventricle, where it is seen as the tail of the caudate nucleus in the adult brain. During the fourth and fifth months the corpus striatum becomes incompletely subdivided by the fibers of the internal capsule into two masses, an inner, the caudate nucleus, and an outer, the lentiform nucleus. In front, the corpus striatum is continuous with the anterior perforated substance; laterally it is confluent for a time with that portion of the wall of the vesicle which is developed into the insula, but this continuity is subsequently interrupted by the fibers of the external capsule. | 28 |
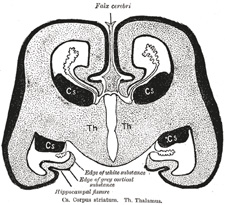 |
FIG. 656– Diagrammatic coronal section of brain to show relations of neopallium. (After His.) Cs. Corpus striatum. Th. Thalamus. (See enlarged image) | | |
| The neopallium (Fig. 656) forms the remaining, and by far the greater, part of the cerebral hemisphere. It consists, at an early stage, of a relatively large, more or less hemispherical cavity—the primitive lateral ventricle—enclosed by a thin wall from which the cortex of the hemisphere is developed. The vesicle expands in all directions, but more especially upward and backward, so that by the third month the hemispheres cover the diencephalon, by the sixth they overlap the mid-brain, and by the eighth the hind-brain. | 29 |
| The median lamina uniting the two hemispheres does not share in their expansion, and thus the hemispheres are separated by a deep cleft, the forerunner of the longitudinal fissure, and this cleft is occupied by a septum of mesodermal tissue which constitutes the primitive falx cerebri. Coincidently with the expansion of the vesicle, its cavity is drawn out into three prolongations which represent the horns of the future lateral ventricle; the hinder end of the vesicle is carried downward and forward and forms the inferior horn; the posterior horn is produced somewhat later, in association with the backward growth of the occipital lobe of the hemisphere. The roof-plate of the primitive fore-brain remains thin and of an epithelial character; it is invaginated into the lateral ventricle along the medial wall of the hemisphere. This invagination constitutes the choroidal fissure, and extends from the interventricular foramen to the posterior end of the vesicle. Mesodermal tissue, continuous with that of the primitive falx cerebri, and carrying bloodvessels with it, spreads between the two layers of the invaginated fold and forms the rudiment of the tela choroidea; the margins of the tela become highly vascular and form the choroid plexuses which for some months almost completely fill the ventricular cavities; the tela at the same time invaginates the epithelial roof of the diencephalon to form the choroid plexuses of the third ventricle. By the downward and forward growth of the posterior end of the vesicle to form the temporal lobe the choroidal fissure finally reaches from the interventricular foramen to the extremity of the inferior horn of the ventricle. | 30 |
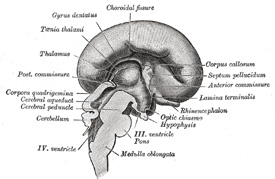 |
FIG. 657– Median sagittal section of brain of human embryo of four months. (Marchand.) (See enlarged image) | | |
| Parallel with but above and in front of the choroidal fissure the medial wall of the cerebral vesicle becomes folded outward and gives rise to the hippocampal fissure on the medial surface and to a corresponding elevation, the hippocampus, within the ventricular cavity. The gray or ganglionic covering of the wall of the vesicle ends at the inferior margin of the fissure is a thickened edge; beneath this the marginal or reticular layer (future white substance) is exposed and its lower thinned edge is continuous with the epithelial invagination covering the choroid plexus (Fig. 656). As a result of the later downward and forward growth of the temporal lobe the hippocampal fissure and the parts associated with it extend from the interventricular foramen to the end of the inferior horn of the ventricle. The thickened edge of gray substance becomes the gyrus dentatus, the fasciola cinerea and the supra- and subcallosal gyri, while the free edge of the white substance forms the fimbria hippocampi and the body and crus of the fornix. The corpus callosum is developed within the arch of the hippocampal fissure, and the upper part of the fissure forms, in the adult brain, the callosal fissure on the medial surface of the hemisphere. | 31 |
| | | The Commissures (Fig. 657).—The development of the posterior commissure has already been referred to (page 743). The great commissures of the hemispheres, viz., the corpus callosum, the fornix, and anterior commissures, arise from the lamina terminalis. About the fourth month a small thickening appears in this lamina, immediately in front of the interventricular foramen. The lower part of this thickening is soon constricted off, and fibers appear in it to form the anterior commissure. The upper part continues to grow with the hemispheres, and is invaded by two sets of fibers. Transverse fibers, extending between the hemispheres, pass into its dorsal part, which is now differentiated as the corpus callosum (in rare cases the corpus callosum is not developed). Into the ventral part longitudinal fibers from the hippocampus pass to the lamina terminalis, and through that structure to the corpora mamillaria; these fibers constitute the fornix. A small portion, lying antero-inferiorly between the corpus callosum and fornix, is not invaded by the commissural fibers; it remains thin, and later a cavity, the cavity of the septum pellucidum, forms in its interior. | 32 |
| | | Fissures and Sulci.—The outer surface of the cerebral hemisphere is at first smooth, but later it exhibits a number of elevations or convolutions, separated from each other by fissures and sulci, most of which make their appearance during the sixth or seventh months of fetal life. The term fissure is applied to such grooves as involve the entire thickness of the cerebral wall, and thus produce corresponding eminences in the ventricular cavity, while the sulci affect only the superficial part of the wall, and therefore leave no impressions in the ventricle. The fissures comprise the choroidal and hippocampal already described, and two others, viz., the calcarine and collateral, which produce the swellings known respectively as the calcar avis and the collateral eminence in the ventricular cavity. Of the sulci the following may be referred to, viz., the central sulcus (fissure of Rolando), which is developed in two parts; the intraparietal sulcus in four parts; and the cingulate sulcus in two or three parts. The lateral cerebral or Sylvian fissure differs from all the other fissures in its mode of development. It appears about the third month as a depression, the Sylvian fossa, on the lateral surface of the hemisphere (Fig. 658); this fossa corresponds with the position of the corpus striatum, and its floor is moulded to form the insula. The intimate connection which exists between the cortex of the insula and the subjacent corpus striatum prevents this part of the hemisphere wall from expanding at the same rate as the portions which surround it. The neighboring parts of the hemisphere therefore gradually grow over and cover in the insula, and constitute the temporal, parietal, frontal, and orbital opercula of the adult brain. The frontal and orbital opercula are the last to form, but by the end of the first year after birth the insula is completely submerged by the approximation of the opercula. The fissures separating the opposed margins of the opercula constitute the composite lateral cerebral fissure. | 33 |
 |
FIG. 658– Outer surface of cerebral hemisphere of human embryo of about five months. (See enlarged image) | | |
| If a section across the wall of the hemisphere about the sixth week be examined microscopically it will be found to consist of a thin marginal or reticular layer, a thick ependymal layer, and a thin intervening mantle layer. Neuroblasts from the ependymal and mantle layers migrate into the deep part of the marginal layer and form the cells of the cerebral cortex. The nerve fibers which form the underlying white substance of the hemispheres consist at first of outgrowths from the cells of the corpora striata and thalami; later the fibers from the cells of the cortex are added. Medullation of these fibers begins about the time of birth and continues until puberty. A summary of the parts derived from the brain vesicles is given in the following table: | 34 |
| Hind-brain or Rhombencephalon |
1. Myelencephalon |
Medulla oblongata |
| Lower part of fourth ventricle. |
|
|
| 2. Metencephalon |
Pons |
| Cerebellum |
| Intermediate part of fourth ventricle. |
|
|
| 3. Isthmus rhombencephali |
Anterior medullary velum |
| Brachia conjunctiva cerebelli. |
| Upper part of fourth ventricle. |
|
|
|
Cerebral peduncles |
| Mid-brain or Mesencephalon………………………… |
Lamina quadrigemina |
|
|
Cerebral aqueduct. |
|
|
|
| Fore-brain or Prosencephalon |
1. Diencephalon |
Thalamus |
| Metathalamus |
| Epithalamus |
| Pars mamillaris hypothalami |
| Posterior part of third ventricle. |
|
|
| 2. Telencephalon |
Anterior part of third ventricle |
| Pars optica hypothalami |
| Cerebral hemispheres |
| Lateral ventricles |
| Interventricular foramen. | |
| | | The Cranial Nerves.—With the exception of the olfactory, optic, and acoustic nerves, which will be especially considered, the cranial nerves are developed in a similar manner to the spinal nerves (see page 735). The sensory or afferent nerves are derived from the cells of the ganglion rudiments of the neural crest. The central processes of these cells grow into the brain and form the roots of the nerves, while the peripheral processes extend outward and constitute their fibers of distribution (Fig. 645). It has been seen, in considering the development of the medulla oblongata (page 739), that the tractus solitarius (Fig. 660), derived from the fibers which grow inward from the ganglion rudiments of the glossopharyngeal and vagus nerves, is the homologue of the oval bundle in the cord which had its origin in the posterior nerve roots. The motor or efferent nerves arise as outgrowths of the neuroblasts situated in the basal laminæ of the mid- and hindbrain. While, however, the spinal motor nerve roots arise in one series from the basal lamina, the cranial motor nerves are grouped into two sets, according as they spring from the medial or lateral parts of the basal lamina. To the former set belong the oculomotor, trochlear, abducent, and hypoglossal nerves; to the latter, the accessory and the motor fibers of the trigeminal, facial, glossopharyngeal, vagus nerves (Figs. 659, 660). | 35 |
 |
FIG. 659– Transverse section of medulla oblongata of human embryo. X 32. (Kollmann) (See enlarged image) | | |
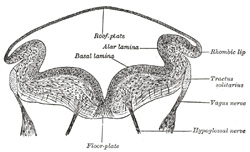 |
FIG. 660– Transverse section of medulla oblongata of human embryo. (After His.) (See enlarged image) | | |
|
|

