| Henry Gray (1821–1865). Anatomy of the Human Body. 1918. |
V. Angiology |
| | | |
| Introduction |
| |
| THE VASCULAR system is divided for descriptive purposes into (a) the blood vascular system, which comprises the heart and bloodvessels for the circulation of the blood; and (b) the lymph vascular system, consisting of lymph glands and lymphatic vessels, through which a colorless fluid, the lymph, circulates. It must be noted, however, that the two systems communicate with each other and are intimately associated developmentally. | 1 |
| The heart is the central organ of the blood vascular system, and consists of a hollow muscle; by its contraction the blood is pumped to all parts of the body through a complicated series of tubes, termed arteries. The arteries undergo enormous ramification in their course throughout the body, and end in minute vessels, called arterioles, which in their turn open into a close-meshed network of microscopic vessels, termed capillaries. After the blood has passed through the capillaries it is collected into a series of larger vessels, called veins, by which it is returned to the heart. The passage of the blood through the heart and blood-vessels constitutes what is termed the circulation of the blood, of which the following is an outline. | 2 |
| The human heart is divided by septa into right and left halves, and each half is further divided into two cavities, an upper termed the atrium and a lower the ventricle. The heart therefore consists of four chambers, two, the right atrium and right ventricle, forming the right half, and two, the left atrium and left ventricle the left half. The right half of the heart contains venous or impure blood; the left, arterial or pure blood. The atria are receiving chambers, and the ventricles distributing ones. From the cavity of the left ventricle the pure blood is carried into a large artery, the aorta, through the numerous branches of which it is distributed to all parts of the body, with the exception of the lungs. In its passage through the capillaries of the body the blood gives up to the tissues the materials necessary for their growth and nourishment, and at the same time receives from the tissues the waste products resulting from their metabolism. In doing so it is changed from arterial into venous blood, which is collected by the veins and through them returned to the right atrium of the heart. From this cavity the impure blood passes into the right ventricle, and is thence conveyed through the pulmonary arteries to the lungs. In the capillaries of the lungs it again becomes arterialized, and is then carried to the left atrium by the pulmonary veins. From the left atrium it passes into the left ventricle, from which the cycle once more begins. | 3 |
| The course of the blood from the left ventricle through the body generally to the right side of the heart constitutes the greater or systemic circulation, while its passage from the right ventricle through the lungs to the left side of the heart is termed the lesser or pulmonary circulation. | 4 |
| It is necessary, however, to state that the blood which circulates through the spleen, pancreas, stomach, small intestine, and the greater part of the large intestine is not returned directly from these organs to the heart, but is conveyed by the portal vein to the liver. In the liver this vein divides, like an artery, and ultimately ends in capillary-like vessels (sinusoids), from which the rootlets of a series of veins, called the hepatic veins, arise; these carry the blood into the inferior vena cava, whence it is conveyed to the right atrium. From this it will be seen that the blood contained in the portal vein passes through two sets of vessels: (1) the capillaries in the spleen, pancreas, stomach, etc., and (2) the sinusoids in the liver. The blood in the portal vein carries certain of the products of digestion: the carbohydrates, which are mostly taken up by the liver cells and stored as glycogen, and the protein products which remain in solution and are carried into the general circulation to the various tissues and organs of the body. | 5 |
| Speaking generally, the arteries may be said to contain pure and the veins impure blood. This is true of the systemic, but not of the pulmonary vessels, since it has been seen that the impure blood is conveyed from the heart to the lungs by the pulmonary arteries, and the pure blood returned from the lungs to the heart by the pulmonary veins. Arteries, therefore, must be defined as vessels which convey blood from the heart, and veins as vessels which return blood to the heart. | 6 |
| | | Structure of Arteries (Fig. 448).—The arteries are composed of three coats: an internal or endothelial coat (tunica intima of Kölliker); a middle or muscular coat (tunica media); and an external or connective-tissue coat (tunica adventitia). The two inner coats together are very easily separated from the external, as by the ordinary operation of tying a ligature around an artery. If a fine string be tied forcibly upon an artery and then taken off, the external coat will be found undivided, but the two inner coats are divided in the track of the ligature and can easily be further dissected from the outer coat. | 7 |
| The inner coat (tunica intima) can be separated from the middle by a little maceration, or it may be stripped off in small pieces; but, on account of its friability, it cannot be separated as a complete membrane. It is a fine, transparent, colorless structure which is highly elastic, and, after death, is commonly corrugated into longitudinal wrinkles. The inner coat consists of: (1) A layer of pavement endothelium, the cells of which are polygonal, oval, or fusiform, and have very distinct round or oval nuclei. This endothelium is brought into view most distinctly by staining with nitrate of silver. (2) A subendothelial layer, consisting of delicate connective tissue with branched cells lying in the interspaces of the tissue; in arteries of less than 2 mm. in diameter the subendothelial layer consists of a single stratum of stellate cells, and the connective tissue is only largely developed in vessels of a considerable size. (3) An elastic or fenestrated layer, which consists of a membrane containing a net-work of elastic fibers, having principally a longitudinal direction, and in which, under the microscope, small elongated apertures or perforations may be seen, giving it a fenestrated appearance. It was therefore called by Henle the fenestrated membrane. This membrane forms the chief thickness of the inner coat, and can be separated into several layers, some of which present the appearance of a net-work of longitudinal elastic fibers, and others a more membranous character, marked by pale lines having a longitudinal direction. In minute arteries the fenestrated membrane is a very thin layer; but in the larger arteries, and especially in the aorta, it has a very considerable thickness. | 8 |
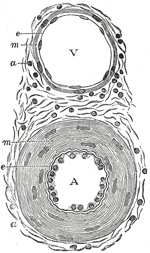 |
FIG. 448– Transverse section through a small artery and vein of the mucous membrane of the epiglottis of a child. X 350. (Klein and Noble Smith.) A. Artery, showing the nucleated endothelium, e, which lines it; the vessel being contracted, the endothelial cells appear very thick. Underneath the endothelium is the wavy elastic lamina. The chief part of the wall of the vessel is occupied by the circular muscle coat m; the rod-shaped nuclei of the muscle cells are well seen. Outside this is a, part of the adventitia. This is composed of bundles of connective tissue fibers, shown in section, with the nuclei of the connective tissue corpuscles. The adventitia gradually merges into the surrounding connective tissue. V. Vein showing a thin endothelial membrane, e, raised accidentally from the intima, which on account of its delicacy is seen as a mere line on the media m. This latter is composed of a few circular unstriped muscle cells a. The adventitia, similar in structure to that of an artery. (See enlarged image) | | |
| The middle coat (tunica media) is distinguished from the inner by its color and by the transverse arrangement of its fibers. In the smaller arteries it consists principally of plain muscle fibers in fine bundles, arranged in lamellæ and disposed circularly around the vessel. These lamellæ vary in number according to the size of the vessel; the smallest arteries having only a single layer (Fig. 449), and those slightly larger three or four layers. It is to this coat that the thickness of the wall of the artery is mainly due (Fig. 448 A, m). In the larger arteries, as the iliac, femoral, and carotid, elastic fibers unite to form lamellæ which alternate with the layers of muscular fibers; these lamellæ are united to one another by elastic fibers which pass between the muscular bundles, and are connected with the fenestrated membrane of the inner coat (Fig. 450). In the largest arteries, as the aorta and innominate, the amount of elastic tissue is very considerable; in these vessels a few bundles of white connective tissue also have been found in the middle coat. The muscle fiber cells are about 50μ in length and contain well-marked, rod-shaped nuclei, which are often slightly curved. | 9 |
| The external coat (tunica adventitia) consists mainly of fine and closely felted bundles of white connective tissue, but also contains elastic fibers in all but the smallest arteries. The elastic tissue is much more abundant next the tunica media, and it is sometimes described as forming here, between the adventitia and media, a special layer, the tunica elastica externa of Henle. This layer is most marked in arteries of medium size. In the largest vessels the external coat is relatively thin; but in small arteries it is of greater proportionate thickness. In the smaller arteries it consists of a single layer of white connective tissue and elastic fibers; while in the smallest arteries, just above the capillaries, the elastic fibers are wanting, and the connective tissue of which the coat is composed becomes more nearly homogeneous the nearer it approaches the capillaries, and is gradually reduced to a thin membranous envelope, which finally disappears. | 10 |
| Some arteries have extremely thin walls in proportion to their size; this is especially the case in those situated in the cavity of the cranium and vertebral canal, the difference depending on the thinness of the external and middle coats. | 11 |
| The arteries, in their distribution throughout the body, are included in thin fibro-areolar investments, which form their sheaths. The vessel is loosely connected with its sheath by delicate areolar tissue; and the sheath usually encloses the accompanying veins, and sometimes a nerve. Some arteries, as those in the cranium, are not included in sheaths. | 12 |
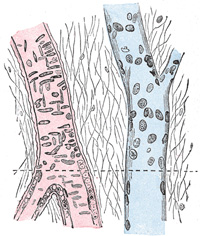 |
FIG. 449– Small artery and vein, pia mater of sheep. X 250. Surface view above the interrupted line; longitudinal section below. Artery in red; vein in blue, (See enlarged image) | | |
| All the larger arteries, like the other organs of the body, are supplied with bloodvessels. These nutrient vessels, called the vasa vasorum, arise from a branch of the artery, or from a neighboring vessel, at some considerable distance from the point at which they are distributed; they ramify in the loose areolar tissue connecting the artery with its sheath, and are distributed to the external coat, but do not, in man, penetrate the other coats; in some of the larger mammals a few vessels have been traced into the middle coat. Minute veins return the blood from these vessels; they empty themselves into the vein or veins accompanying the artery. Lymphatic vessels are also present in the outer coat. | 13 |
| Arteries are also supplied with nerves, which are derived from the sympathetic, but may pass through the cerebrospinal nerves. They form intricate plexuses upon the surfaces of the larger trunks, and run along the smaller arteries as single filaments, or bundles of filaments which twist around the vessel and unite with each other in a plexiform manner. The branches derived from these plexuses penetrate the external coat and are distributed principally to the muscular tissue of the middle coat, and thus regulate, by causing the contraction and relaxation of this tissue the amount of blood sent to any part. | 14 |
| | | The Capillaries.—The smaller arterial branches (excepting those of the cavernous structure of the sexual organs, of the splenic pulp, and of the placenta) terminate in net-works of vessels which pervade nearly every tissue of the body. These vessels, from their minute size, are termed capillaries. They are interposed between the smallest branches of the arteries and the commencing veins, constituting a net-work, the branches of which maintain the same diameter throughout; the meshes of the net-work are more uniform in shape and size than those formed by the anastomoses of the small arteries and veins. | 15 |
| The diameters of the capillaries vary in the different tissues of the body, the usual size being about 8μ. The smallest are those of the brain and the mucous membrane of the intestines; and the largest those of the skin and the marrow of bone, where they are stated to be as large as 20μ in diameter. The form of the capillary net varies in the different tissues, the meshes being generally rounded or elongated. | 16 |
| The rounded form of mesh is most common, and prevails where there is a dense network, as in the lungs, in most glands and mucous membranes, and in the cutis; the meshes are not of an absolutely circular outline, but more or less angular, sometimes nearly quadrangular, or polygonal, or more often irregular. | 17 |
| Elongated meshes are observed in the muscles and nerves, the meshes resembling parallelograms in form, the long axis of the mesh running parallel with the long axis of the nerve or muscle. Sometimes the capillaries have a looped arrangement; a single vessel projecting from the common net-work and returning after forming one or more loops, as in the papillæ of the tongue and skin. | 18 |
| The number of the capillaries and the size of the meshes determine the degree of vascularity of a part. The closest network and the smallest interspaces are found in the lungs and in the choroid coat of the eye. In these situations the interspaces are smaller than the capillary vessels themselves. In the intertubular plexus of the kidney, in the conjunctiva, and in the cutis, the interspaces are from three to four times as large as the capillaries which form them; and in the brain from eight to ten times as large as the capillaries in their long diameters, and from four to six times as large in their transverse diameters. In the adventitia of arteries the width of the meshes is ten times that of the capillary vessels. As a general rule, the more active the function of the organ, the closer is its capillary net and the larger its supply of blood; the meshes of the network are very narrow in all growing parts, in the glands, and in the mucous membranes, wider in bones and ligaments which are comparatively inactive; bloodvessels are nearly altogether absent in tendons, in which very little organic change occurs after their formation. In the liver the capillaries take a more or less radial course toward the intralobular vein, and their walls are incomplete, so that the blood comes into direct contact with the liver cells. These vessels in the liver are not true capillaries but “sinusoids;” they are developed by the growth of columns of liver cells into the blood spaces of the embryonic organ. | 19 |
 |
FIG. 450– Section of a medium-sized artery. (After Grünstein.) (See enlarged image) | | |
| Structure.—The wall of a capillary consists of a fine transparent endothelial layer, composed of cells joined edge to edge by an interstitial cement substance, and continuous with the endothelial cells which line the arteries and veins. When stained with nitrate of silver the edges which bound the epithelial cells are brought into view (Fig. 451). These cells are of large size and of an irregular polygonal or lanceolate shape, each containing an oval nucleus which may be displayed by carmine or hematoxylin. Between their edges, at various points of their meeting, roundish dark spots are sometimes seen, which have been described as stomata, though they are closed by intercellular substance. They have been believed to be the situations through which the colorless corpuscles of the blood, when migrating from the bloodvessels, emerge; but this view, though probable, is not universally accepted. | 20 |
| Kolossow describes these cells as having a rather more complex structure. He states that each consists of two parts: of hyaline ground plates, and of a protoplasmic granular part, in which is imbedded the nucleus, on the outside of the ground plates. The hyaline internal coat of the capillaries does not form a complete membrane, but consists of “plates” which are inelastic, and though in contact with each other are not continuous; when therefore the capillaries are subjected to intravascular pressure, the plates become separated from each other; the protoplasmic portions of the cells, on the other hand, are united together. In some organs, e. g., the glomeruli of the kidneys, intercellular cement cannot be demonstrated in the capillary wall and the cells are believed to form a syncytium. | 21 |
| In many situations a delicate sheath or envelope of branched nucleated connective tissue cells is found around the simple capillary tube, particularly in the larger ones; and in other places, especially in the glands, the capillaries are invested with retiform connective tissue. | 22 |
| | | Sinusoids.—In certain organs, viz., the heart, the liver, the suprarenal and parathyroid glands, the glomus caroticum and glomus coccygeum, the smallest bloodvessels present various differences from true capillaries. They are wider, with an irregular lumen, and have no connective tissue covering, their endothelial cells being in direct contact with the cells of the organ. Moreover, they are either arterial or venous and not intermediate as are the true capillaries. These vessels have been called sinusoids by Minot. They are formed by columns of cells or trabeculæ pushing their way into a large bloodvessel or blood space and carrying its endothelium before them; at the same time the wall of the vessel or space grows out between the cell columns. | 23 |
| | | Structure of Veins.—The veins, like the arteries, are composed of three coats: internal, middle, and external; and these coats are, with the necessary modifications, analogous to the coats of the arteries; the internal being the endothelial, the middle the muscular, and the external the connective tissue or areolar (Fig. 452). The main difference between the veins and the arteries is in the comparative weakness of the middle coat in the former. | 24 |
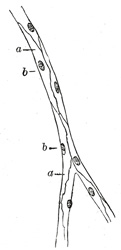 |
FIG. 451– Capillaries from the mesentery of a guinea-pig, after treatment with solution of nitrate of silver. a. Cells. b. Their nuclei. (See enlarged image) | | |
| In the smallest veins the three coats are hardly to be distinguished (Fig. 449). The endothelium is supported on a membrane separable into two layers, the outer of which is the thicker, and consists of a delicate, nucleated membrane (adventitia), while the inner is composed of a network of longitudinal elastic fibers (media). In the veins next above these in size (0.4 mm. in diameter), according to Kölliker, a connective tissue layer containing numerous muscle fibers circularly disposed can be traced, forming the middle coat, while the elastic and connective tissue elements of the outer coat become more distinctly perceptible. In the middle-sized veins the typical structure of these vessels becomes clear. The endothelium is of the same character as in the arteries, but its cells are more oval and less fusiform. It is supported by a connective tissue layer, consisting of a delicate net-work of branched cells, and external to this is a layer of elastic fibers disposed in the form of a net-work in place of the definite fenestrated membrane seen in the arteries. This constitutes the internal coat. The middle coat is composed of a thick layer of connective tissue with elastic fibers, intermixed, in some veins, with a transverse layer of muscular tissue. The white fibrous element is in considerable excess, and the elastic fibers are in much smaller proportion in the veins than in the arteries. The outer coat consists, as in the arteries, of areolar tissue, with longitudinal elastic fibers. In the largest veins the outer coat is from two to five times thicker than the middle coat, and contains a large number of longitudinal muscular fibers. These are most distinct in the inferior vena cava, especially at the termination of this vein in the heart, in the trunks of the hepatic veins, in all the large trunks of the portal vein, and in the external iliac, renal, and azygos veins. In the renal and portal veins they extend through the whole thickness of the outer coat, but in the other veins mentioned a layer of connective and elastic tissue is found external to the muscular fibers. All the large veins which open into the heart are covered for a short distance with a layer of striped muscular tissue continued on to them from the heart. Muscular tissue is wanting: (1) in the veins of the maternal part of the placenta; (2) in the venous sinuses of the dura mater and the veins of the pia mater of the brain and medulla spinalis; (3) in the veins of the retina; (4) in the veins of the cancellous tissue of bones; (5) in the venous spaces of the corpora cavernosa. The veins of the above-mentioned parts consist of an internal endothelial lining supported on one or more layers of areolar tissue. | 25 |
| Most veins are provided with valves which serve to prevent the reflux of the blood. Each valve is formed by a reduplication of the inner coat, strengthened by connective tissue and elastic fibers, and is covered on both surfaces with endothelium, the arrangement of which differs on the two surfaces. On the surface of the valve next the wall of the vein the cells are arranged transversely; while on the other surface, over which the current of blood flows, the cells are arranged longitudinally in the direction of the current. Most commonly two such valves are found placed opposite one another, more especially in the smaller veins or in the larger trunks at the point where they are joined by smaller branches; occasionally there are three and sometimes only one. The valves are semilunar. They are attached by their convex edges to the wall of the vein; the concave margins are free, directed in the course of the venous current, and lie in close apposition with the wall of the vein as long as the current of blood takes its natural course; if, however, any regurgitation takes place, the valves become distended, their opposed edges are brought into contact, and the current is interrupted. The wall of the vein on the cardiac side of the point of attachment of each valve is expanded into a pouch or sinus, which gives to the vessel, when injected or distended with blood, a knotted appearance. The valves are very numerous in the veins of the extremities, especially of the lower extremities, these vessels having to conduct the blood against the force of gravity. They are absent in the very small veins, i. e., those less than 2 mm. in diameter, also in the venæ cavæ, hepatic, renal, uterine, and ovarian veins. A few valves are found in each spermatic vein, and one also at its point of junction with the renal vein or inferior vena cava respectively. The cerebral and spinal veins, the veins of the cancellated tissue of bone, the pulmonary veins, and the umbilical vein and its branches, are also destitute of valves. A few valves are occasionally found in the azygos and intercostal veins. Rudimentary valves are found in the tributaries of the portal venous system. | 26 |
 |
FIG. 452– Section of a medium-sized vein. (See enlarged image) | | |
| The veins, like the arteries, are supplied with nutrient vessels, vasa vasorum. Nerves also are distributed to them in the same manner as to the arteries, but in much less abundance. | 27 |
|
|

